科研│澳门科大&广州中医药大学:多组学分析揭示了疏肝消脂汤保护机体免受代谢功能障碍相关性脂肪性肝炎的损害的机制
- 2026-06-26 11:28:11
 点击蓝字“代谢组metabolome”,轻松关注不迷路
点击蓝字“代谢组metabolome”,轻松关注不迷路

生科云网址:https://www.bioincloud.tech/
编译:微科盟 梅子,编辑:微科盟X、江舜尧。
微科盟原创微文,欢迎转发转载。
背景:代谢功能障碍相关性脂肪性肝炎(MASH)是一种威胁人类健康的肝脏疾病,目前治疗策略有限。作为治疗MASH的临床中药复方,舒肝消脂(SGXZ)汤具有明确疗效,但其治疗MASH的机制尚不十分清楚。
目的:通过多组学分析和动物实验验证,探索疏肝消脂(SGXZ)汤治疗MASH的潜在机制。
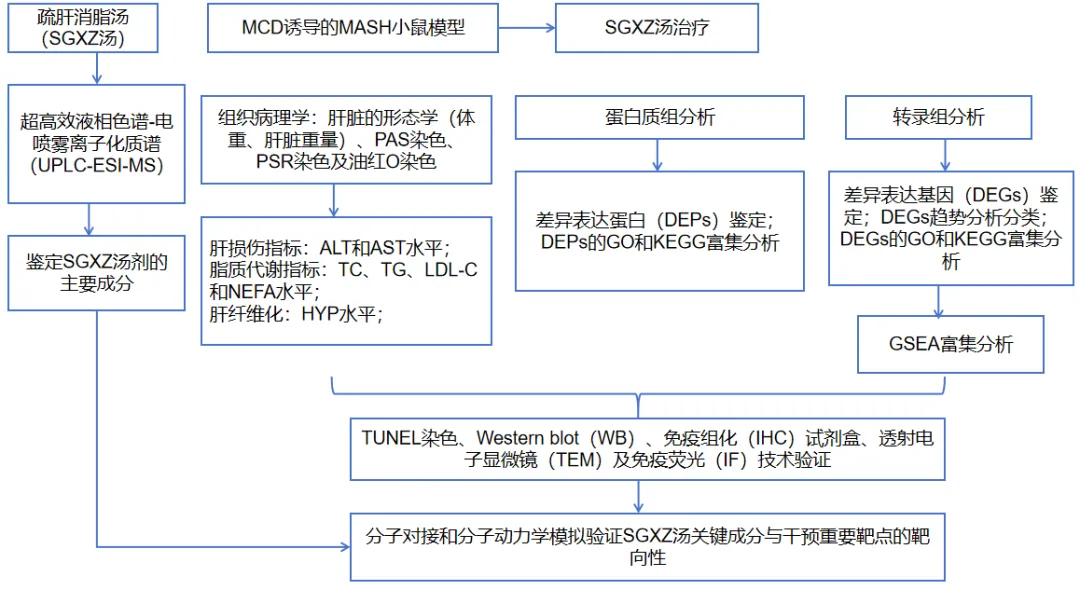
方法:采用超高效液相色谱-电喷雾离子化质谱(UPLC-ESI-MS)方法鉴定SGXZ汤的主要成分。通过过碘酸-雪夫反应(PAS)、苦味酸天狼星红(PSR)及油红O染色评估SGXZ汤对MCD诱导的MASH小鼠模型的治疗效果。采用多组学分析SGXZ汤对MASH的作用机制。通过TUNEL染色、Western blot(WB)、免疫组化(IHC)试剂盒、透射电子显微镜(TEM)及免疫荧光(IF)技术验证SGXZ汤对MASH的作用机制。最后,通过分子对接和分子动力学模拟验证了SGXZ汤关键成分与干预重要靶点的靶向性。
结果:通过UPLC-ESI-MS分析,研究者从SGXZ汤中获得了30种主要活性成分。SGXZ汤改善了MASH,这体现在组织病理学、肝功能指标、脂质和纤维化指标的改善上。蛋白质组学和转录组学结果均提示铁死亡在SGXZ 汤干预MASH 中的重要作用,与铁死亡相关的通路是这些分析中获得的主要显著通路。此外,SGXZ 汤治疗减少了细胞死亡、炎症和氧化应激水平,并恢复了 MCD 诱导的 MASH 小鼠受损的线粒体形态。此外,机制实验证明,SGXZ汤治疗改善了 MASH 小鼠的铁代谢和脂质过氧化失衡,并激活了 Xc-系统。
结论:SGXZ 汤对 MASH 确实具有治疗作用,其机制可能与其调节 p53/SLC7A11/GPX4 通路以减少铁死亡有关。
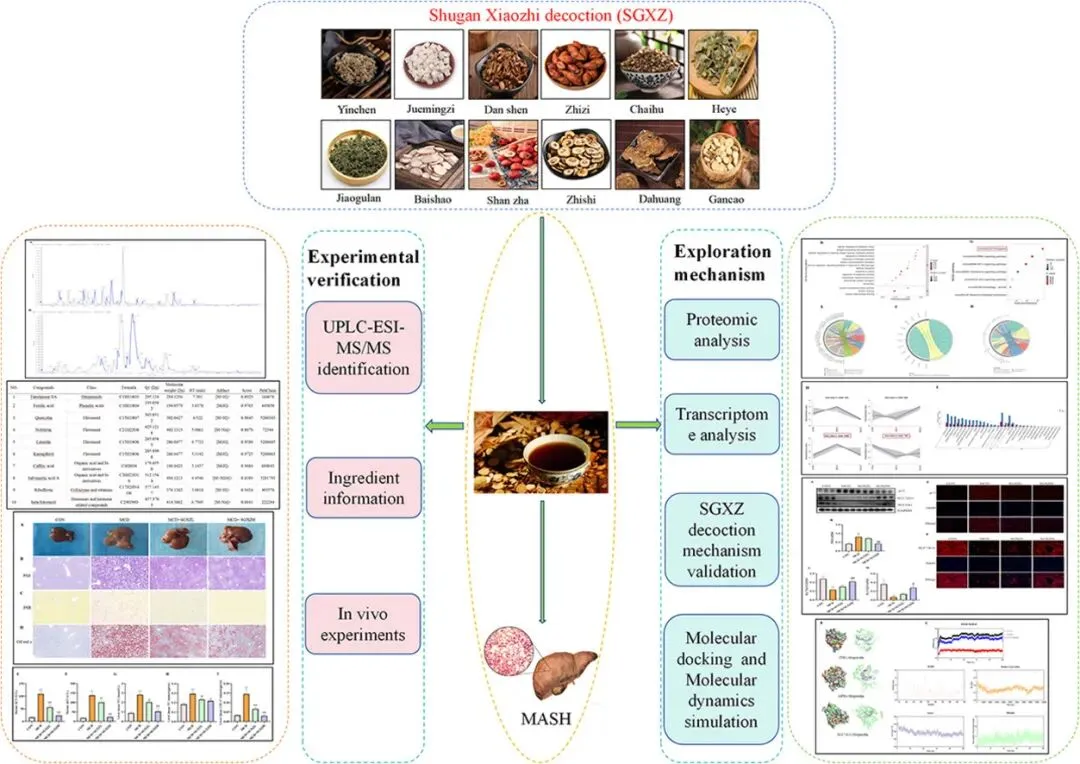
论文ID
实验设计
结果
1 采用UPLC-ESIMS/MS鉴别SGXZ汤的主要成分
研究显示,SGXZ 汤(表1)在负离子模式和正离子模式下的基峰色谱图(BPC)结果如图 1A、B 所示,SGXZ汤的总离子色谱图(TIC)结果见补充图1A、B。研究者在SGXZ汤中鉴定出30种主要成分,各成分的详细信息见表2。这些结果初步揭示了SGXZ汤的主要成分。
表1.SGXZ汤的英文名、拉丁名、中文名及药材克数。
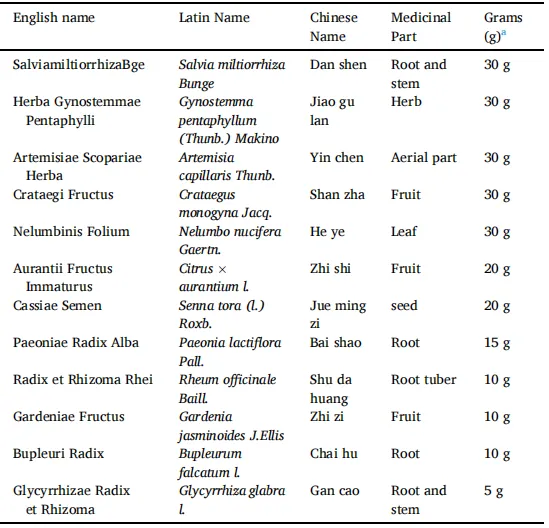
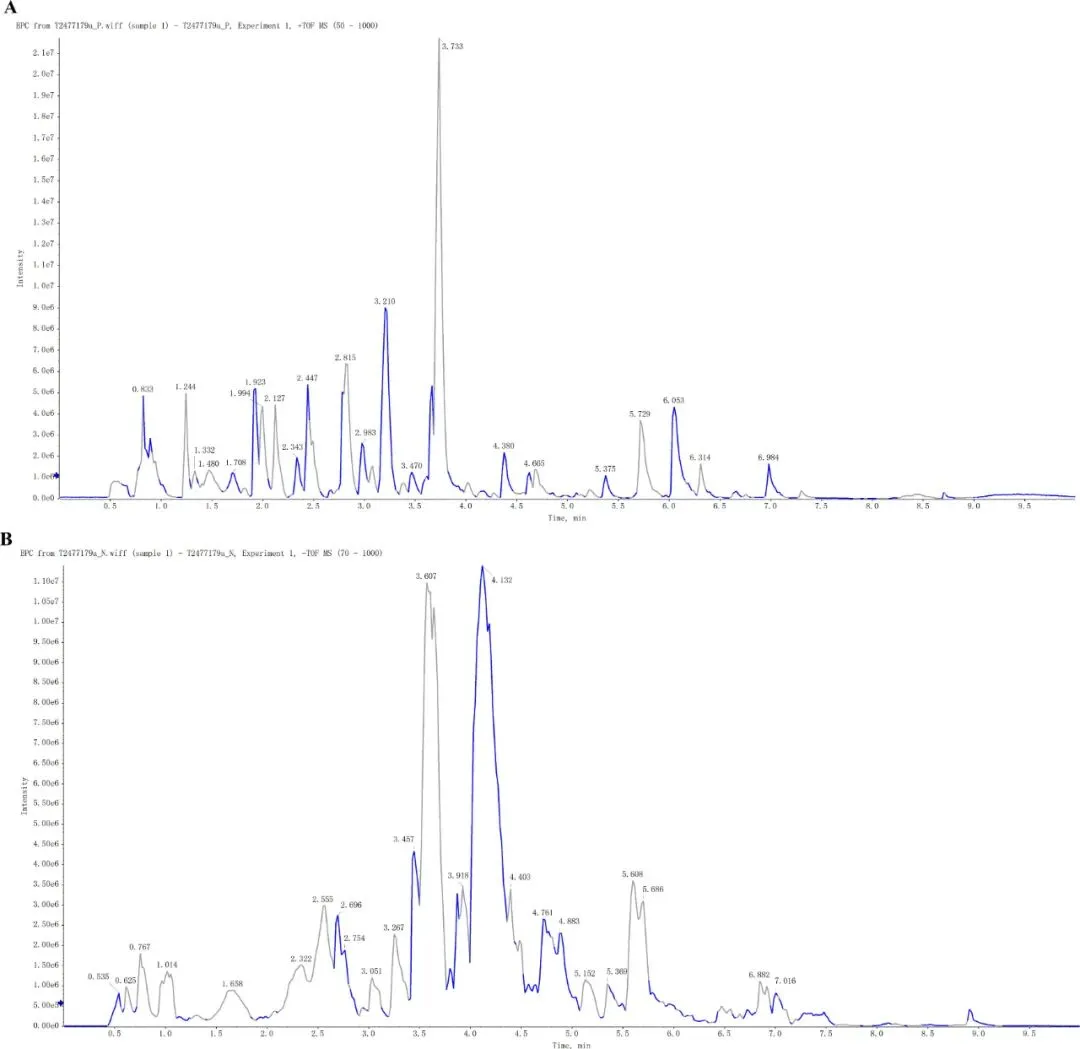
图1.SGXZ汤在正负离子模式下采用UPLC-ESI-MS检测的总基峰色谱图(BPC)。(A) SGXZ汤在正离子模式下的色谱图。(B) SGXZ汤在负离子模式下的色谱图。
表2通过UPLC-ESI-MS法鉴别SGXZ汤的化学成分。
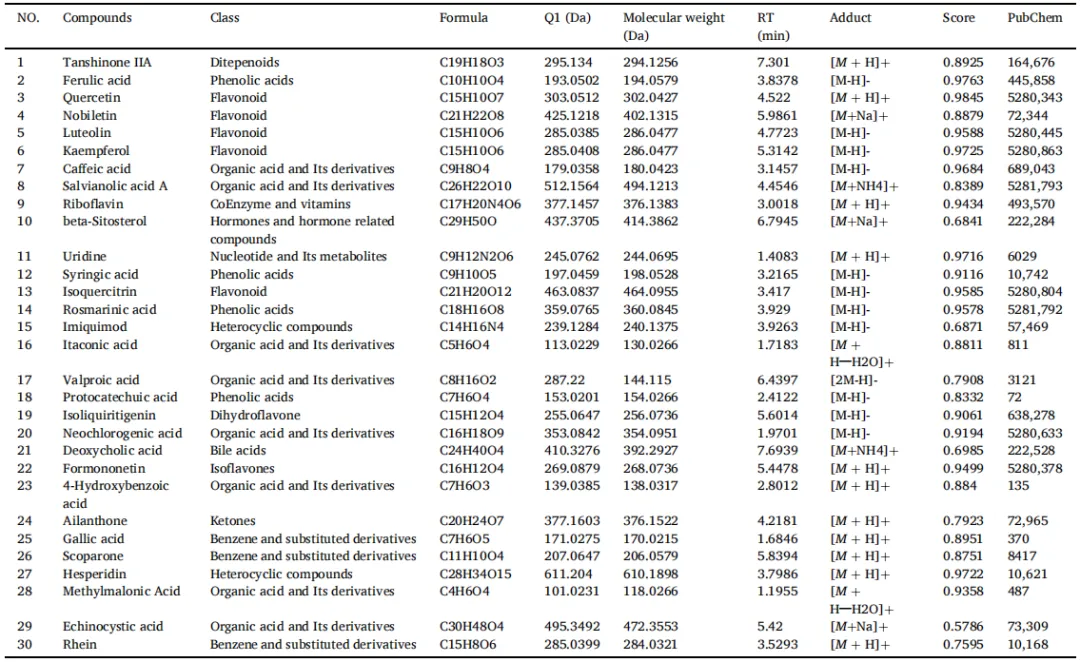
2 SGXZ汤减轻MCD诱导的MASH模型肝脏病理损伤,保护肝功能
为了探讨SGXZ汤对MASH的治疗效果,研究人员通过MCD饮食诱导小鼠MASH模型,并随后用SGXZ汤进行治疗。根据肝脏的形态学特征,CON组的肝脏表面光滑,呈深红色,肝叶边缘锋利; 喂食MCD饮食6周后,与CON组相比,MCD组的肝脏体积增加,肝脏颜色变浅;然而,与MCD组相比,SGXZ汤各剂量组的肝脏颜色略微发红,且各组的肝脏形态得到改善(图2A)。与正常饮食组小鼠相比,MCD喂养小鼠的肝脏出现了典型的MASH病理特征,如广泛的脂肪变性、肝细胞肿胀改变、炎症细胞浸润(图2B)、肝纤维化(图2C)、肝脂质蓄积(图2D),但SGXZ汤改善了这些病理改变。为确定SGXZ汤对MCD诱导的肝损伤的保护作用,研究人员测定了ALT和AST水平。如图2E、F所示,与CON组相比,MCD组的ALT和AST水平显著升高(p<0.001)。然而,SGXZ汤干预后,AST和ALT水平显著降低(p<0.01)。此外,为进一步探索SGXZ汤对MCD诱导的肝脂质蓄积和纤维化的治疗效果,研究人员检测了肝组织中TG、TC、LDL-C、NEFA和HYP的水平。同样,SGXZ汤治疗抑制了MCD在MASH小鼠中诱导的这些指标的上调(p<0.001)(图2G-K)。此外,SGXZ 汤治疗降低了MCD 诱导的 MASH 小鼠的体重、肝脏重量和肝脏指数水平(p<0.01)(图 2l-N)。综上所述,研究者得出结论,SGXZ 汤改善了MCD 诱导的小鼠 MASH。
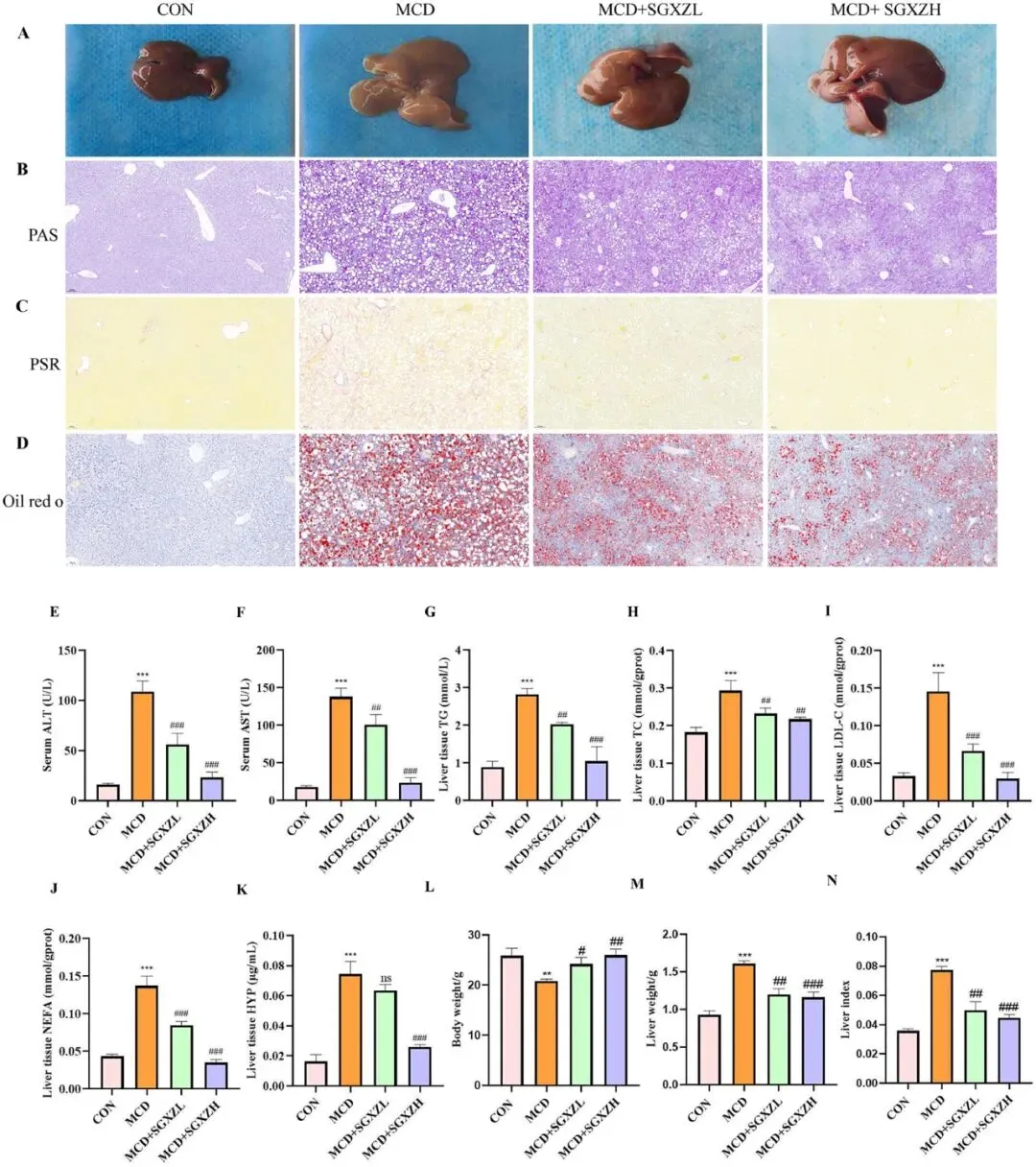
图2. SGXZ汤可减轻MCD诱导的MASH模型肝脏病理损伤并保护肝功能。(A-D)展示了肝脏形态学、PAS染色、PSR染色及油红O染色的代表性图像,比例尺= 50μm。(E-F)各组肝损伤指标ALT和AST水平的柱状图。(G-J)各组脂质代谢指标TC、TG、LDL-C和NEFA水平的柱状图。(K)各组肝纤维化HYP水平的柱状图。(l-N)各组体重、肝脏重量及肝功能指数的柱状图。**p<0.01,***p<0.001与对照组相比。#p<0.05,##p<0.01,###p<0.01与MCD组相比(n = 8)。
3 SGXZ汤干预后MCD诱导MASH的蛋白质组学变化
为探究疏肝消脂方(SGXZ)治疗代谢功能障碍相关性脂肪性肝炎(MASH)的新机制,研究人员对各组肝组织进行了4D无标记定量蛋白质组学分析。韦恩图结果显示,研究者在MCD(蛋氨酸-胆碱缺乏饮食)模型组 vs 空白对照组(CON),以及SGXZ治疗组 vs MCD组之间,共筛选出1026种差异表达蛋白(DEPs),其中包括超氧化物歧化酶1(SOD1)和溶质载体家族3成员2(SLC3A2)。火山图分析进一步表明:与CON组相比,MCD组中有961种蛋白上调、685种蛋白下调;而与MCD组相比,SGXZ治疗组中有606种蛋白上调、721种蛋白下调(图3B,C)。值得注意的是,SOD1和SLC3A2的蛋白水平在MCD组中显著下调,经SGXZ干预后则明显回升。在获得DEPs后,研究者通过GO/KEGG功能注释分析发现:GO条目主要富集于细胞对氧化应激的反应、过氧化氢应答、自噬调控及凋亡过程调节等生物学过程(图3D-F)。KEGG通路分析显示(图3G,H),SGXZ干预组与对照组、模型组相比,显著富集的通路包括铁死亡(Ferroptosis)、过氧化物酶体增殖物激活受体通路(PPAR Pathway)、缺氧诱导因子-1通路(HIF-1 Pathway)以及p53信号通路。这些结果提示,SGXZ方对MASH的治疗作用可能与其调控铁死亡机制密切相关。
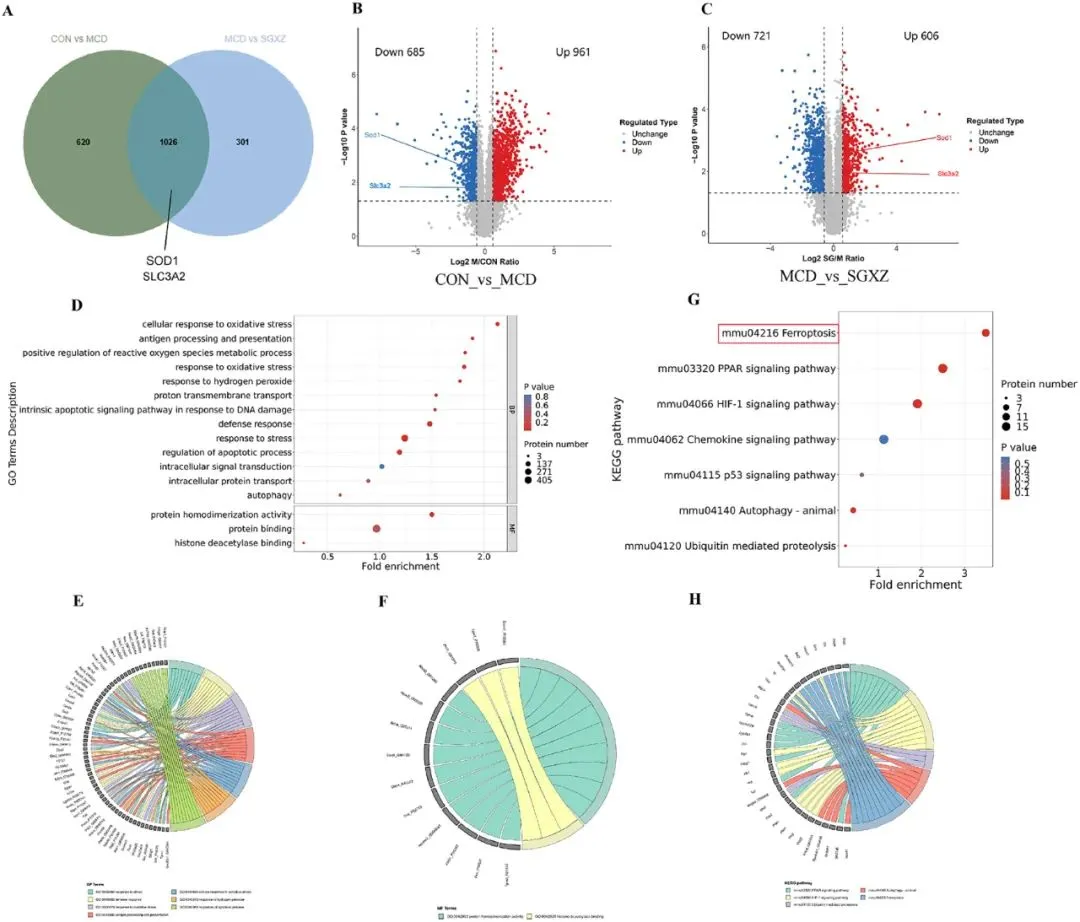
图3.SGXZ汤干预后MCD诱导的MASH模型中蛋白质组学变化。(A)三组差异表达蛋白(DEPs)的重叠情况。(B-C)对照组、模型组与SGXZ组差异表达蛋白的火山图,红色和蓝色圆点分别表示显著上调和下调的DEPs。(D-F)MCD组与SGXZ组差异表达蛋白的GO富集分析以气泡图和弦图形式展示,蛋白质被分类到不同的生物过程(E)分子功能(F),气泡大小表示属于特定通路的蛋白质数量,颜色代表p值。(G-H)MCD组与SGXZ组差异表达蛋白的KEGG富集分析。
4 SGXZ汤干预MCD诱导-MASH的转录分析
为了进一步探讨SGXZ汤对MASH的机制,研究人员对接受SGXZ汤治疗的MASH模型进行了转录组分析。二维主成分分析图(图4A)显示,对照组(CON)、SGXZ汤组和MCD组之间存在显著的分离趋势。三维主成分分析图(图4B)显示,SGXZ汤组有向对照组倾斜的趋势,但仍与MCD组明显分离。随后,热图结果清晰地显示了不同组的转录谱(图4C)。研究者设置p<0.05和|Log2FC| > 1的阈值以筛选并获得差异表达基因(DEGs)。在CON vs MCD的比较中,鉴定出2651个上调和1071个下调的DEGs。MCD vs SGXZ的比较中,发现了1095个上调和2841个下调的DEGs(图4D-F)。韦恩图显示,MCD vs CON以及SGXZ vs MCD之间有2598个重叠的DEGs(图4G)。为探讨SGXZ汤干预后MCD诱导的MASH模型中的转录组变化,研究者对DEGs进行了趋势分析并分为四类。在第3类中,MCD组的基因表达水平上调,而CON组和SGXZ汤组则下调; 在第4类中,MCD组的基因表达水平下调,而CON组和SGXZ汤组的基因表达水平上调(图4H)。在MCD vs SGXZ汤组的比较中,下调的基因集中在GO术语中,如细胞过程、生物调节、生物过程调节、含蛋白质的复合物、结合和抗氧化活性; MCD组vs SGXZ汤组比较中上调的基因集中在GO术语如免疫系统过程、催化活性、生物过程的正调节和对刺激的反应(图4I)。其中,细胞过程和结合类基因受影响最大。为了找到可能受影响的通路,研究者获得了KEGG富集圆图(图4J)和气泡图(图4K)。KEGG富集包括HIF-1通路、过氧化物酶体、PPAR通路、p53通路、铁死亡、氧化磷酸化、线粒体自噬-动物、PI3K-AKT和MAPK等通路,其中PI3K-AKT和MAPK通路受影响最大。这些结果还表明,SGXZ 汤干扰 MASH 的机制可能与其对铁死亡的调节有关,这与蛋白质组学结果一致。
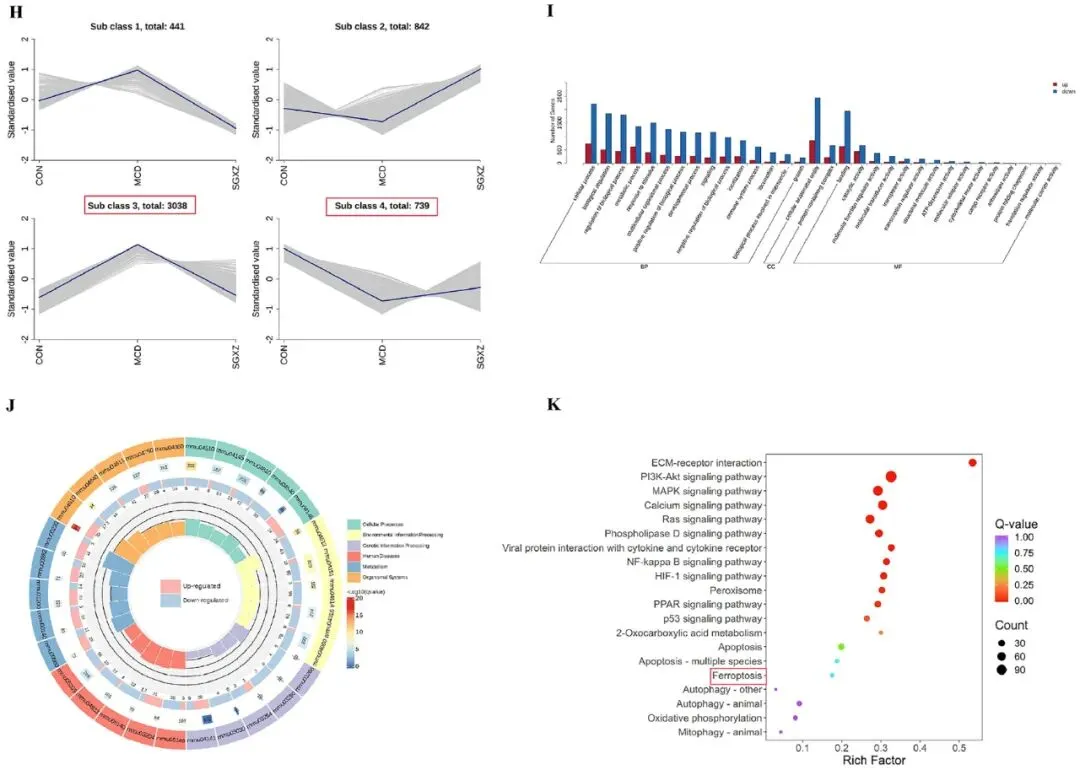
图4.SGXZ汤干预MCD诱导-MASH的转录组分析。(A-B)各组二维和三维PCA分布趋势图。(C)全基因热力图。(D-E)对照组、模型组与SGXZ汤组差异基因的MA图谱。(F)差异基因统计图谱。(G)对照组、模型组与SGXZ汤组差异基因维恩图。(H)差异表达基因K均值聚类图。(I)MCD组与SGXZ汤组差异基因的GO富集分析。(J)MCD组与SGXZ汤组KEGG富集环图。(K)MCD组与SGXZ汤组差异表达基因的KEGG富集分析。
5 基于SGXZ汤干预MCD诱导-MASH转录组学的GSEA富集分析
GSEA富集分析结果显示,与对照组(CON)相比,MCD组在氧化磷酸化、HIF-1通路、泛素介导的蛋白水解和mTOR通路中存在负调控(图5A)。相反,在SGXZ汤与模型组的比较中,氧化磷酸化、HIF-1通路、泛素介导的蛋白水解以及mTOR通路显示出正向调控(图5B)。GSEA富集的KEGG富集图谱包括氧化磷酸化、HIF-1通路、泛素介导的蛋白水解和mTOR通路(图5C)。这些结果表明,SGXZ汤可能通过干扰与氧化应激相关的通路来改善MASH。
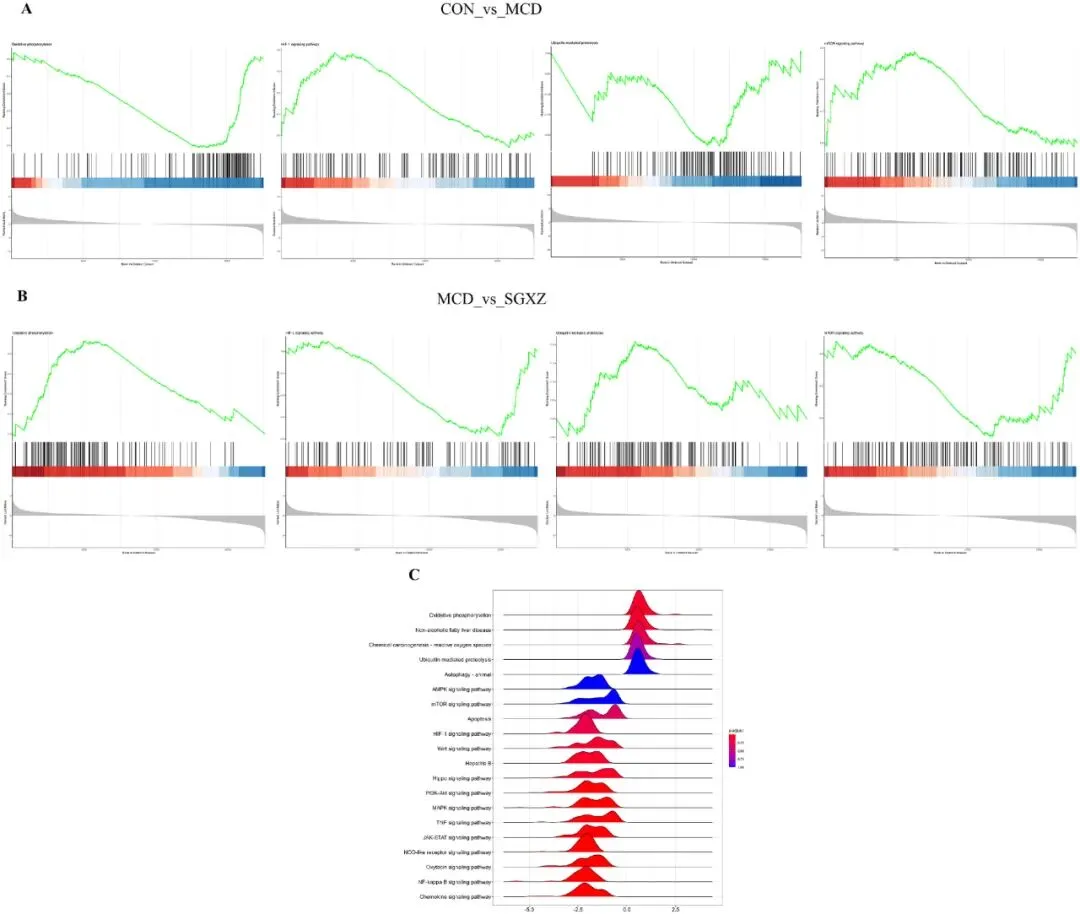
图5.基于SGXZ汤干预MCD诱导-MASH的转录组学的GSEA富集分析。(A-B)对照组、MCD组和SGXZ汤组的KEGG数据库GESA富集通路图谱。(C)MCD组和SGXZ汤组的KEGG数据库GESA富集脊线图谱。
6 SGXZ汤降低MCD诱导的MASH模型中肝细胞死亡和炎症反应
铁死亡是一种依赖于铁的细胞死亡形式。铁死亡及其引发的炎症反应在MASH的发病机制中起着重要作用。因此,为了验证蛋白质组学和转录组学结果,研究者观察了MASH小鼠肝脏在SGXZ汤干预前后细胞死亡和炎症水平的变化。TUNEL染色显示,与对照组相比,MCD组检测到大量细胞死亡,然而,SGXZ汤治疗能够减少细胞死亡(p<0.01)(图6A、B)。此外,研究者测量了SGXZ汤干预前后与炎症相关的蛋白质CD68、CD80、F4/80和HMGB1的水平。WB结果表明,SGXZ汤治疗抑制了MASH小鼠中CD68、CD80、F4/80和HMGB1水平的升高(p<0.001)(图6C-G)。免疫组化(IHC)结果也显示,SGXZ汤干预抑制了MASH小鼠中CD80、CD68和HMGB1水平的上调(p<0.05)(图6H-K)。这些结果表明,SGXZ汤在MASH模型中具有抗炎和减少肝细胞死亡的作用。
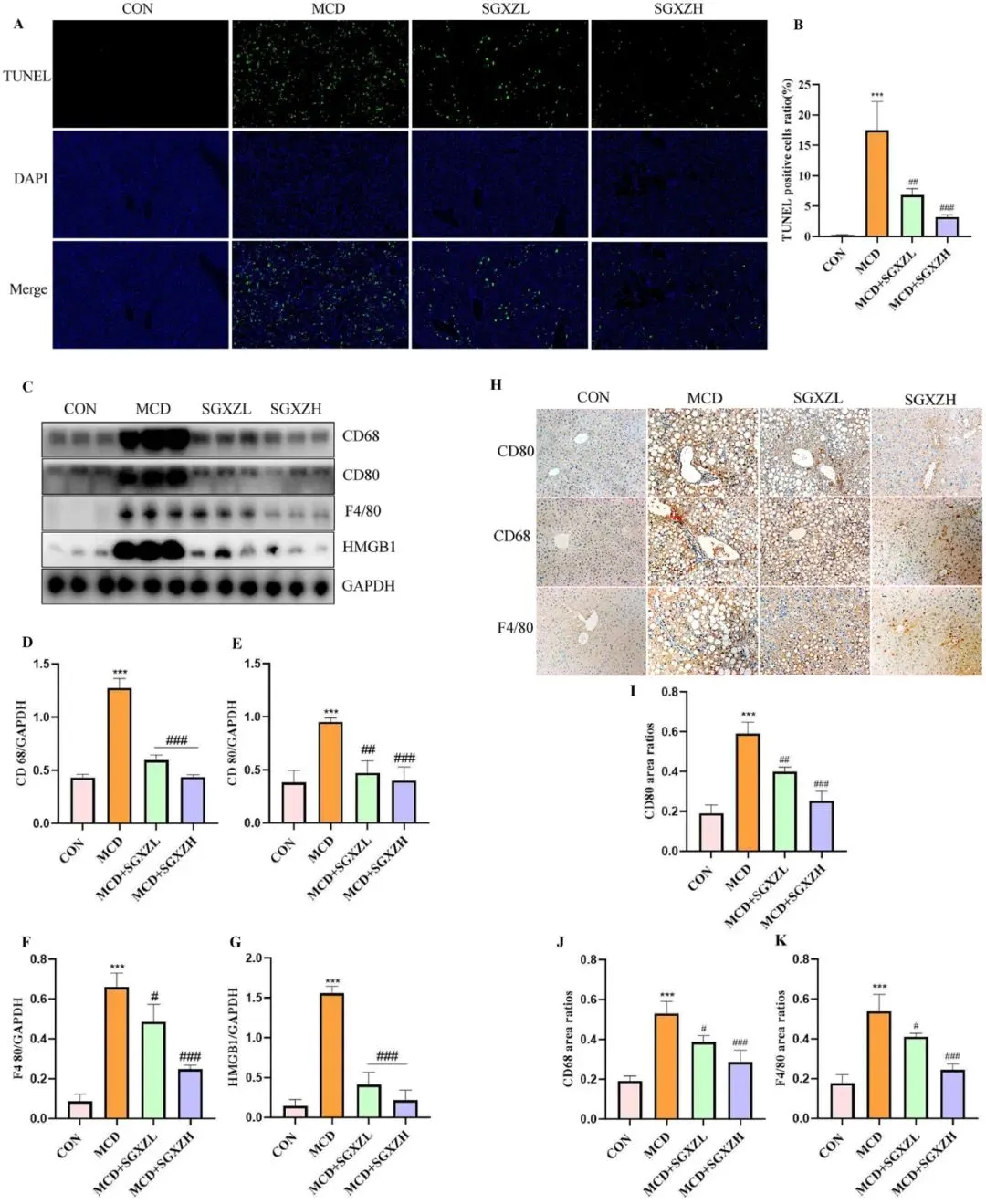
图6. SGXZ汤降低MCD诱导的MASH模型中肝细胞死亡及炎症反应。(A)TUNEL染色显示的肝细胞死亡代表性图像,比例尺= 200 μm。(B)各组肝细胞死亡数量统计。(C)肝脏CD68、CD80、F4/80及HMGB1蛋白的典型Western blot结果。(D-G)CD68、CD80、F4/80及HMGB1蛋白表达量的光密度分析(以GAPDH含量为内参)。(H)肝脏CD68、CD80和F4/80蛋白的免疫组化典型图像,比例尺= 50μm。(I-K)CD68、CD80和F4/80蛋白表达量的免疫组化定量分析。AOD=IOD/面积。***p<0.001(与对照组相比);#p<0.05;##p<0.01;###p<0.01(与MCD组相比,n=8)。
7 SGXZ汤缓解MCD诱导的MASH模型中肝脏氧化应激,保护线粒体结构
铁死亡的本质是线粒体形态的改变以及细胞膜磷脂因过量脂质过氧化物引起的过氧化损伤。脂质过氧化物的积累导致的氧化应激既是铁死亡的特征,也是MASH进展的重要致病因素。因此,为了进一步验证蛋白质组学和转录组学结果,研究者检测了MASH小鼠在SGXZ汤干预前后与氧化应激相关的蛋白质CAT和SOD1的水平。Western blot结果显示,与对照组相比,MCD组的CAT和SOD1水平显著下调(p<0.001),而SGXZ汤治疗抑制了MCD诱导的CAT和SOD1水平下调(p<0.05)(图7A-C),这些结果在图7F,G中进一步得到证实 。此外,免疫荧光(IF)结果也显示,SGXZ 汤干预逆转了MCD 诱导的 MASH 小鼠 CAT 和 SOD1 水平的下调(图7H、I)。 此外,SGXZ 汤还抑制了MCD 诱导的 MDA 水平上调和 GSH 水平下降(p<0.05)(图 7D、E)。此外,透射电子显微镜结果表明,CON组线粒体的形态和大小正常,线粒体嵴清晰可见。在MCD组,线粒体嵴破裂,线粒体密度增加,线粒体变小且肿胀。这些情况在SGXZ汤治疗后得到缓解(图7J)。这些结果表明,SGXZ 汤能够改善氧化应激并保护MASH 模型中的线粒体,这与蛋白质组学和转录组学分析的结果一致,表明 SGXZ 汤能够通过抑制铁死亡来改善MASH。
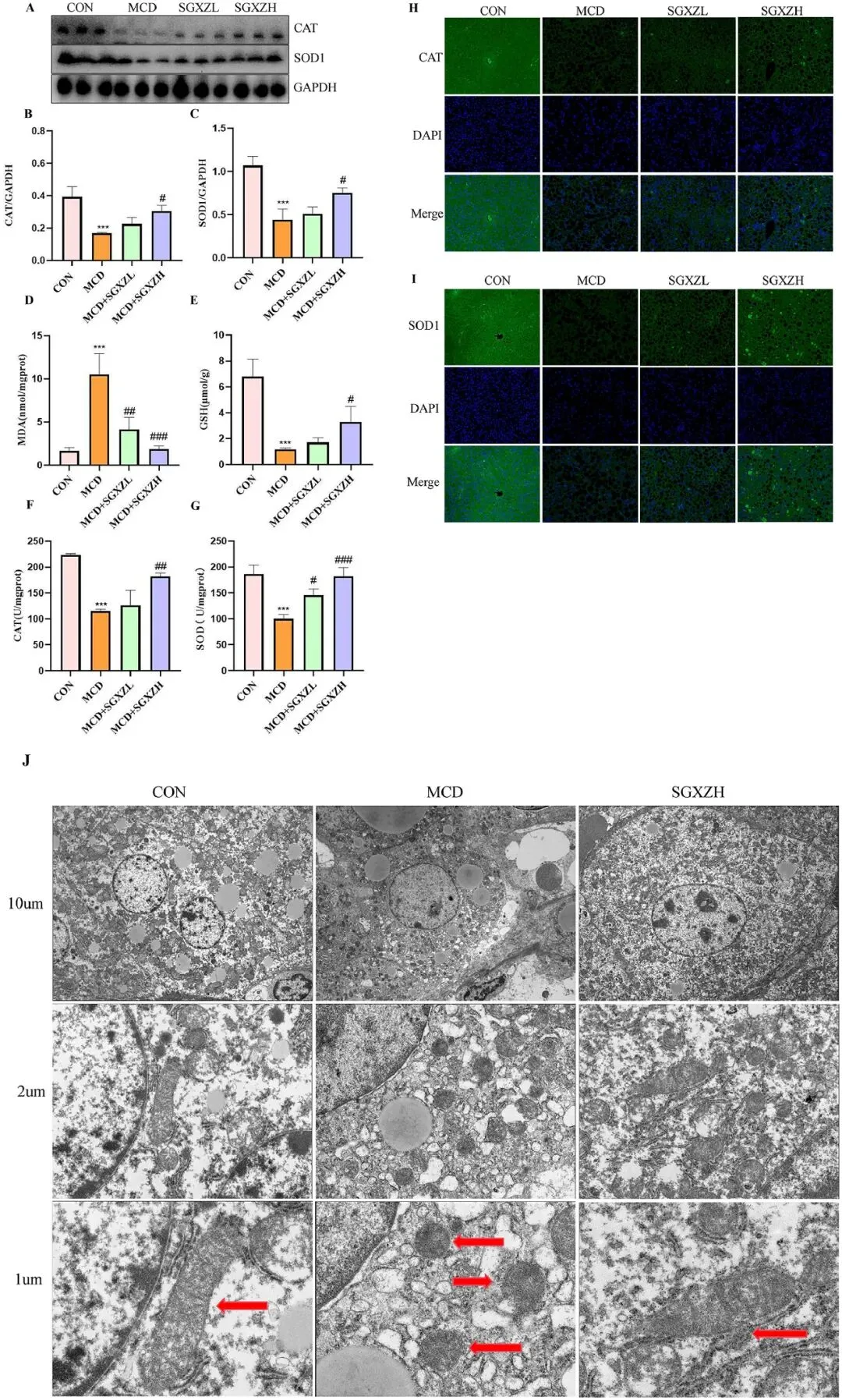
图7. SGXZ汤缓解MCD诱导的MASH模型中肝脏氧化应激并维持线粒体结构。(A)肝脏CAT和SOD1蛋白的代表性Western blot图像。(B-C)CAT和SOD1蛋白表达量与GAPDH含量标准化后的光密度分析结果。(D-G)各组间丙二醛(MDA)、超氧化物歧化酶(SOD)、谷胱甘肽(GSH)和CAT水平的柱状图。(H-I)肝脏CAT和SOD1蛋白的代表性免疫荧光图像。比例尺= 50 μm。(J)各组线粒体超微结构的代表性透射电镜图像。比例尺= 10 μm、2 μm、1 μm。***p<0.001 vs.对照组;#p<0.05,##p<0.01,###p<0.01 vs. MCD组(n = 8)。
8 SGXZ汤减轻MCD诱导的MASH模型中铁代谢失衡
为了探讨SGXZ汤通过抑制铁死亡改善MASH的机制,研究者进行了以下实验。铁代谢失衡、脂质过氧化以及Xc系统失衡是铁死亡发生的主要机制。铁代谢失衡会触发MASH中的铁死亡。因此,调节铁代谢可帮助预防铁死亡的发生。与CON组相比,MCD组中与铁转运相关的蛋白质TFR1和DMT1表达上调(p<0.001),但经SGXZ汤治疗后其表达下调(图8A-C、F)。此外,SGXZ 汤干预可抑制MCD 诱导的 MASH 小鼠中铁蛋白(FTH1、FTL)水平的下调(p<0.05)(图 8D、E、G)。这些结果表明,SGXZ 汤可通过纠正MASH 模型中铁代谢紊乱来抑制铁死亡。
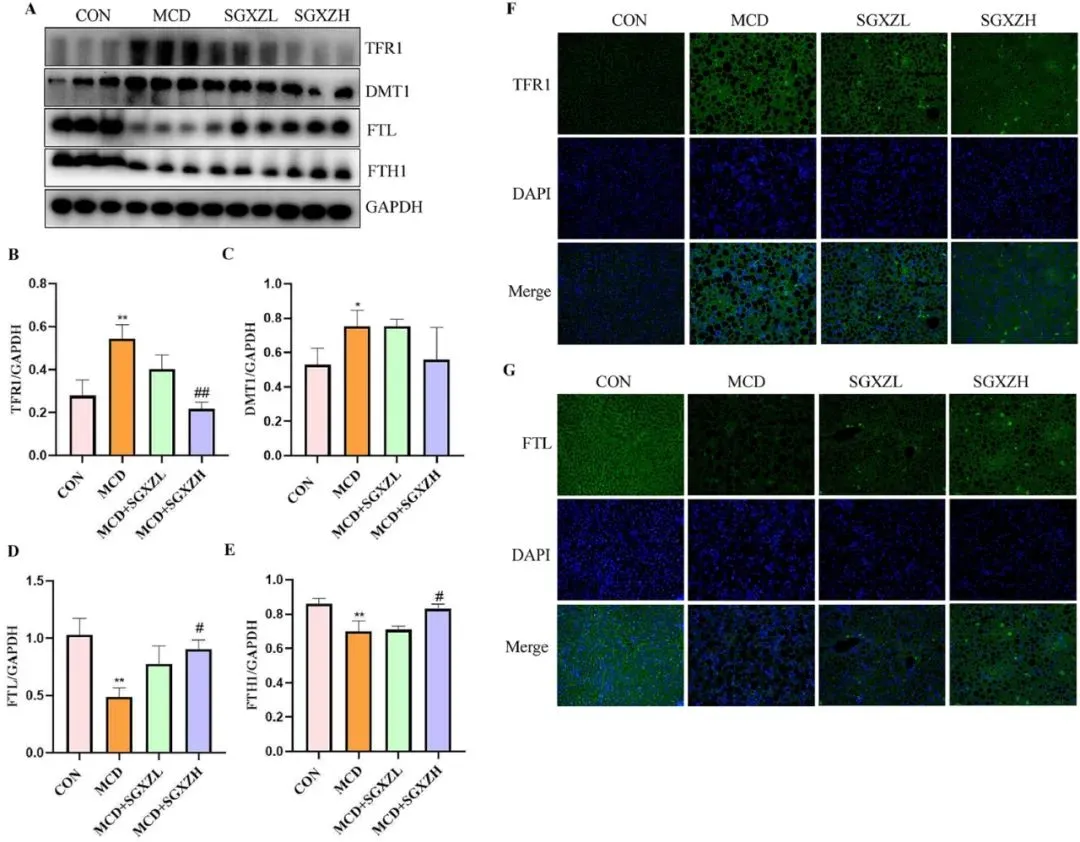
图8. SGXZ汤缓解MCD诱导的MASH模型中铁代谢失衡。(A)肝脏中TFR1、DMT1、FTL和FTH1蛋白的代表性Western blot图像。(B-E)以GAPDH含量为内参的TFR1、DMT1、FTL和FTH1蛋白表达量光密度分析结果。(F-G)肝脏中TFR1和FTL蛋白的代表性免疫荧光图像。比例尺= 50 μm。***p<0.001(与对照组相比);#p<0.05,##p<0.01,###p<0.01(与MCD组相比,n=8)。
9 SGXZ汤抑制MCD诱导的MASH模型脂质过氧化
为了验证SGXZ汤通过抑制铁死亡改善MASH的机制,研究者进行了进一步的实验验证。铁死亡伴随着大量脂质过氧化物的释放,而脂质过氧化是MASH的重要病理因素,两者具有共同的病理基础。因此,研究者还探讨了SGXZ汤治疗是否会影响脂质过氧化关键酶的水平。与对照组相比,MCD诱导的MASH模型中COX2和ACSL4的表达上调,而GPX4的表达下调(p<0.001)。然而,SGXZ 汤治疗可逆转这些关键酶的表达(p<0.05)(图 9A-G)。这些结果表明,SGXZ 汤可通过阻断MASH 模型中的脂质过氧化来抑制铁死亡。
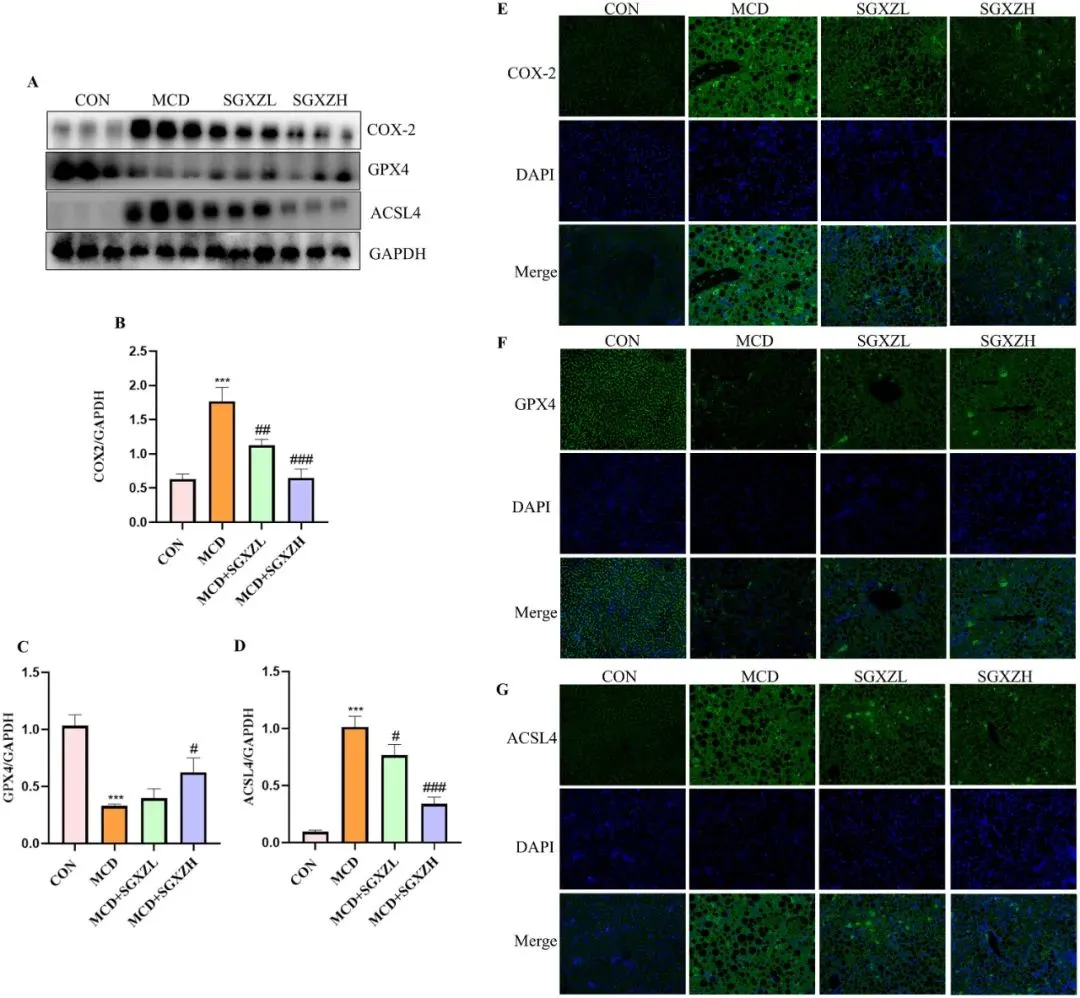
图9. SGXZ汤抑制MCD诱导的MASH模型脂质过氧化。(A)肝脏中COX2、GPX4和ACSL4蛋白的代表性Western blot图像。(B-D)以GAPDH含量为内参的COX2、GPX4和ACSL4蛋白表达量光密度分析结果。(E-G)肝脏中COX2、GPX4和ACSL4蛋白的代表性免疫荧光图像。比例尺= 50 μm。***p<0.001 vs.对照组;#p<0.05,##p<0.01,###p<0.001 vs. MCD组(n = 8)。
10 SGXZ汤对MCD诱导的MASH模型中p53和Xc-系统的表达有影响
为了进一步验证SGXZ汤通过抑制铁死亡改善MASH的机制,研究者进行了进一步的验证。由Xc-系统失衡引起的铁死亡也可促进MASH的发生发展。Xc-系统是一个包含SLC7A11和SLC3A2的抗转运体,负责摄取半胱氨酸并外排谷氨酸。抑制该抗转运体可能阻碍谷胱甘肽(GSH)的合成并导致铁死亡。作为铁死亡相关SLC7A11抑制剂的p53可影响半胱氨酸摄取和GSH合成,进而引发脂质过氧化和铁死亡。因此,在MCD诱导的MASH模型中,研究者检测了SGXZ汤干预前后p53的表达水平。Western blot结果表明,MASH小鼠的p53水平上调,而SGXZ汤治疗逆转了MCD诱导的p53表达水平变化(p<0.01)(图10A、B)。免疫荧光(IF)也被用于测量p53表达,结果也表明SGXZ汤抑制了MCD诱导的p53表达上调(图10E)。此外,Western Blot(WB)和/或免疫荧光(IF)结果均表明,SGXZ 汤可通过抑制MCD 诱导的 SLC7A11 和 SLC3A2 水平下调来激活 Xc-系统(图10C-F)。综上所述,这些结果表明,SGXZ 汤可能通过调节p53/SLC7A11/GPX4 通路在 MASH 模型中抑制铁死亡。
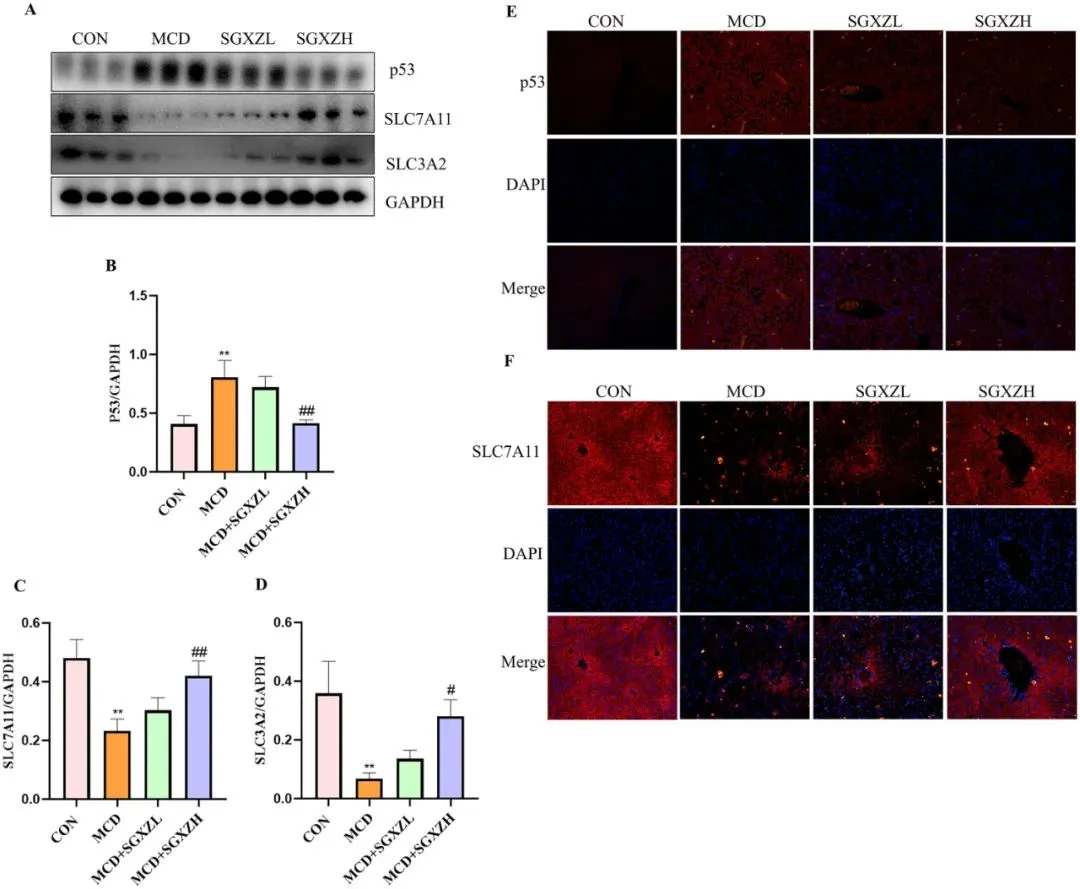
图10. SGXZ汤对MCD诱导的MASH模型中p53及Xc-系统表达的影响。(A)肝脏中p53、SLC7A11和SLC3A2蛋白的代表性Western blot图像。(B-D)经GAPDH含量标准化后的p53、SLC7A11和SLC3A2蛋白表达量光密度分析结果。(E-F)肝脏中p53和SLC7A11蛋白的代表性免疫荧光图像。比例尺= 50μm。**p<0.01 vs.对照组;#p<0.05,##p<0.01 vs. MCD组(n = 8)。
11 分子对接和分子动力学模拟结果
为了验证SGXZ汤活性成分与铁死亡靶点的结合活性,研究者对SGXZ汤的30种主要活性成分与核心铁死亡靶点(TFR1、GPX4、SLC7A11)进行了批量分子对接(图11A)。SGXZ 汤的活性成分与铁死亡核心蛋白中结合能较高的结合对进行了展示,其中SLC7A11 与橙皮苷的结合亲和力最高,为 -10.4,优于其他基因(图 11B,补充图 2)。此外,表面等离子体共振(SPR)结果也显示,橙皮苷与GPX4、SLC7A11和TFR1蛋白具有良好的亲和力(补充图3)。为了验证SLC7A11与橙皮苷的分子对接模拟,研究者进行了分子动力学模拟。SLC7A11-橙皮苷复合物的RMSD曲线在20 ns后达到平衡,平均RMSD值为0.5 nm。RMSF分析识别出SLC7A11和橙皮苷中的关键柔性区域,其中200至250位点和300至325位点具有更大的残基柔性。Rg值在80 ns内保持稳定,随后在2.25至2.3 nm之间波动。相应的SASA值保持在200 nm²左右稳定。在100 ns的模拟过程中,SLC7A11-Hesperidin复合物中的氢键数量最多可达11个(图11C)。上述结果表明,小分子能够与蛋白质形成稳定的复合物并发挥重要作用。
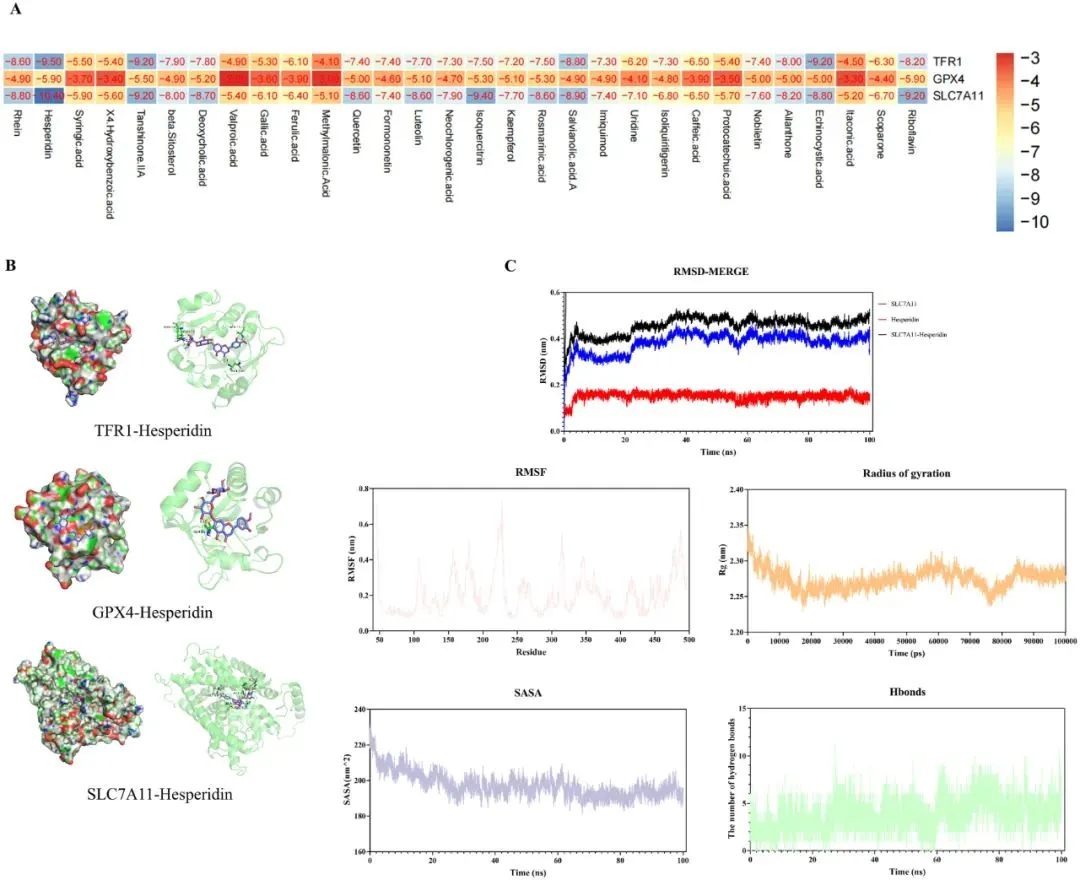
图11.分子对接与分子动力学模拟结果。(A)SGXZ汤中30种活性成分与关键靶点的对接得分热力图。(B)关键靶点与化合物的典型对接复合物示意图。(C)SLC7A11-橙皮苷复合物的分子动力学模拟分析,包含RMSD曲线、RMSF曲线、Rg曲线、SASA曲线及氢键分析。
讨论
铁死亡在MASH的发生发展中发挥着关键作用。铁死亡依赖于铁离子,当氧化还原系统因过量脂质过氧化物积累而失衡时,铁死亡会被诱导。肝脏中铁水平的上调可能诱发MASLD,而过量铁水平可能通过增加炎症和纤维化,从而将MASLD转化为MASH。脂质积累可能导致肝脏脂肪变性,过量脂质积累会引发氧化应激。 而氧化应激在MASH的进展中发挥关键作用。因此,铁死亡和MASH共享许多相同的病理因素。研究表明,西洛他唑通过减轻肝铁过载介导的铁死亡来缓解MASH的进展。研究表明,ENO3通过调节GPX4表达和脂质积累来缓解铁死亡,从而改善MASH。槲皮素通过减轻铁死亡来缓解脂质积累。因此,铁死亡是MASH中重要的病理机制和治疗靶点。
目前,中医在MASH的预防和治疗中发挥着积极作用。黄连-五味子药对通过抑制NLRP3炎症小体缓解MASH。四物汤通过调节ACSL4介导的铁死亡来改善MASH。SGXZ汤作为一种经典复方,已临床应用超过十年,在治疗MASH方面具有优势。临床研究表明,SGXZ 汤与多烯磷脂酰胆碱联合治疗MASH 的疗效显著优于多烯磷脂酰胆碱单药治疗,尤其在改善肝功能和脂质方面。研究者之前的基础研究也表明,SGXZ 汤可通过其抗炎和抗凋亡作用改善MASH。此外,研究者小组的先前研究还表明,SGXZ 汤可通过调节线粒体功能和氧化应激来减轻MASH。然而,尚不清楚SGXZ汤是否能通过抑制铁死亡来改善MASH。在本研究中,研究者探讨了SGXZ汤缓解MASH的机制,并重点关注其对铁死亡的调节作用,铁死亡是MASH中一个重要的致病因素。
在本研究中,经过SGXZ汤治疗后,MASH小鼠的炎症浸润、脂肪变性、气球样变、脂质沉积和纤维化均得到抑制。此外,SGXZ 汤改善了肝脏形态、肝脏重量和肝功能。这些结果表明,SGXZ 汤在改善MASH 表型方面有效,证实了 SGXZ 汤对MASH 的改善作用,这与研究者的初步发现一致。为探索SGXZ汤改善MASH的分子机制,本研究采用蛋白质组学和转录组学技术,分析了SGXZ汤对MASH相关分子机制的影响。蛋白质组学结果显示,差异表达蛋白主要富集于氧化应激响应通路。转录组学结果也表明,与抗氧化相关的基因表达明显下调。同时,蛋白质组学和转录组学结果均表明,差异表达的蛋白质或基因与铁死亡相关。而氧化应激是铁死亡的核心触发因素,氧化应激诱导的脂质过氧化也是铁死亡的标志性特征。研究者的结果显示,SGXZ 汤干预上调了氧化应激相关蛋白CAT、SOD1 和 GSH 的表达,并抑制了 MCD 诱导的 MASH 模型中 MDA 蛋白的表达,这些结果与蛋白质组学结果一致。而GSH 耗竭是铁死亡的关键生化特征。此外,SGXZ 汤治疗还恢复了MASH 小鼠受损线粒体的形态,这种线粒体损伤属于铁死亡的特殊形态学特征。此外,SGXZ 汤干预减轻了MASH 小鼠的细胞死亡并抑制了炎症反应,这进一步表明 MASH 模型中存在铁死亡,这些结果还表明 SGXZ 汤可能通过靶向铁死亡来改善MASH。
为了探讨SGXZ汤通过靶向铁死亡机制缓解MASH的药理学机制,研究者分析了蛋白质组学和转录组学研究的KEGG通路结果。这些结果表明,铁死亡及其相关通路是SGXZ汤干预MASH的关键通路。研究表明,铁死亡参与了MASH的发生发展,可能主要与铁代谢紊乱、脂质过氧化及Xc系统失衡引起的铁死亡相关。因此,通过调节脂质过氧化、纠正铁代谢及Xc系统失衡来抑制铁死亡,可能是改善MASH的主要机制。
为了验证SGXZ汤通过靶向铁死亡机制缓解MASH的机制,研究者观察了MASH小鼠在SGXZ汤干预前后铁死亡相关铁代谢的变化。铁代谢主要由肝脏调节,肝脏通过分泌铁代谢调节因子维持铁稳态。铁稳态调节因子主要包括铁转运蛋白、铁调节蛋白和铁储存蛋白。TFR1、DMT1、铁蛋白(FTH1和FTL)以及铁蛋白1等蛋白质在维持全身铁稳态中发挥重要作用。有研究提出,MASLD中肝铁含量升高可能与转运蛋白TFRs和DMT1的上调有关。抑制DMT1和TFR的表达可促进肝细胞铁排泄,从而改善MASLD。 此外,研究还表明,MASH患者肠道中DMT1和TFR1 mRNA的表达在慢性炎症发作时会上调。铁蛋白由两种亚单位以不同比例组成,即FTH和FTL,它们是重要的铁相关因子。研究表明,Caveolin-1 可通过激活 FTL/FTH 通路改善 MASLD。在本研究中,MCD 诱导的 MASH 模型中 TFR1 和 DMT1 的水平上调,而 FTH 和 FTL 的表达下调,这与上述发现一致。然而,SGXZ 汤治疗逆转了 MCD 治疗引起的这些病理状态。这些结果表明,SGXZ 汤可能通过改善铁死亡过程中的铁代谢紊乱来改善 MASH。
接下来,研究者进一步测量了MASH小鼠在SGXZ汤干预前后铁死亡过程中的脂质过氧化水平。脂质过氧化是铁死亡的主要特征之一,也是驱动其发生的重要因素之一。脂质氧化酶可分为三类:环氧化酶(COXs)、细胞色素P450(CYPs)和脂氧合酶(LOXs),其中LOXs对铁死亡尤为重要。LOXs 在正常状态下诱导脂肪酸过氧化物的产生,而 GPX4 将脂质过氧化物转化为无毒脂质并防止铁死亡。ACSL4是脂质代谢的调节因子,可抑制GPX4,进而引发铁死亡。因此,ACSL4的缺失可能通过限制氧化敏感性脂肪酸的积累来抑制铁死亡。COX2是铁死亡的已知生物标志物。同样,在本研究中,研究者发现SGXZ汤干预抑制了MASH小鼠中ACSL4和COX2水平的升高,并上调了GPX4水平,这表明SGXZ汤对MASH的改善作用可能与其减轻脂质过氧化和因此抑制铁死亡的能力有关。
Xc–GSH-GPX4 轴是关键的抗氧化系统,可诱导谷胱甘肽(GSH)合成并抑制脂质过氧化,从而减少铁死亡。Xc-系统是铁死亡的重要组成部分,激活 Xc 系统轴有助于预防铁死亡的发生,这对改善 MASH 具有重要意义。 这与本研究结果相似,研究者的研究也表明,SGXZ 汤在 MASH 小鼠模型中激活了 Xc 系统并抑制了 p53 的表达。这些结果表明,SGXZ 汤通过调节 p53/SLC7A11/GPX4 通路减少铁死亡,从而缓解了 MASH。
但本研究也存在一些局限性:首先,未测定SGXZ汤在体内的血药浓度;其次,需要通过体外实验进一步深入探究SGXZ汤治疗MASH的机制;最后,需解决SGXZ汤的临床转化性、药代动力学问题及监管挑战等。
综上所述,为了深入理解SGXZ汤治疗MASH的机制,研究者采用了蛋白质组学和转录组学分析的结合方法,初步预测了SGXZ汤的调节机制。因此,研究者在MASH小鼠模型中进行了实验,并证实SGXZ汤改善了与MASH相关的生理参数。通过整合蛋白质组学与转录组学联合分析,本研究阐明了SGXZ汤抗MASH作用的机制,该机制可能通过调节p53/SLC7A11/GPX4通路从而缓解铁死亡(图12)。这些结果有助于研究者更好地认识SGXZ汤的治疗潜力,并为开发新的MASH治疗方法提供启示。
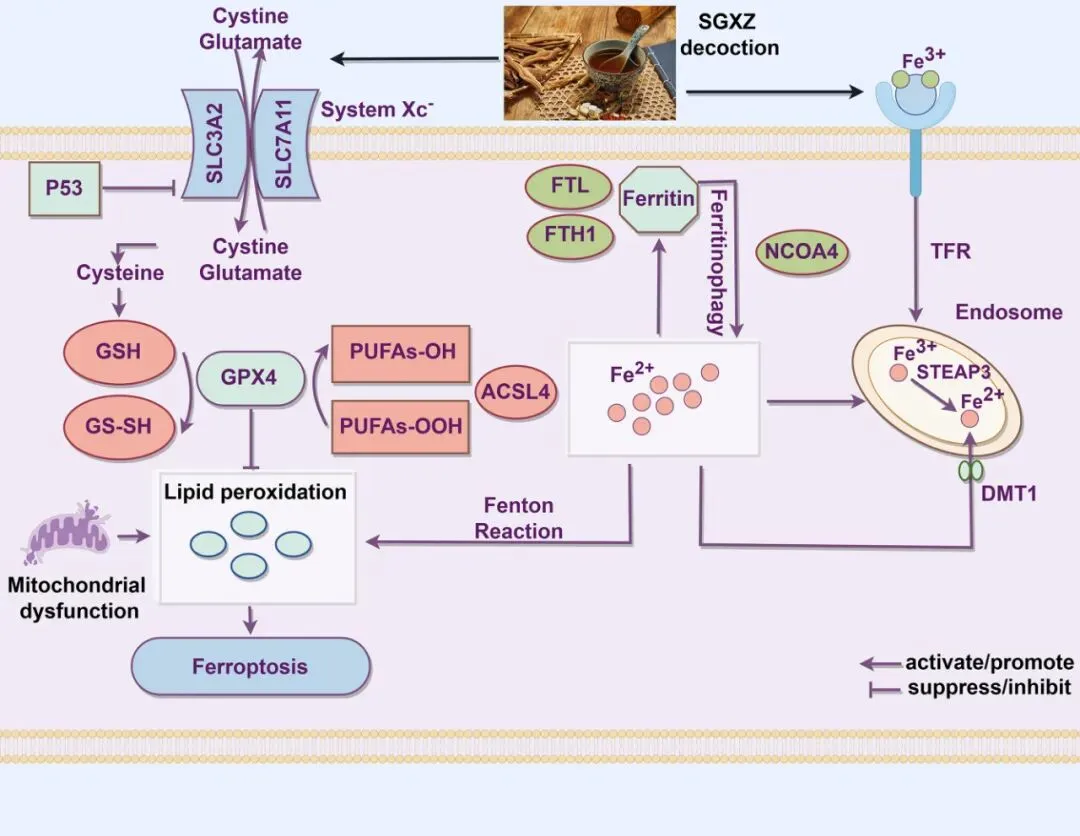
图12.SGXZ汤通过抑制铁死亡改善MASH的机制。
结论
简而言之,铁死亡在MASH的发生发展中起着关键作用。在本研究中,SGXZ汤通过调节p53/SLC7A11/GPX4通路抑制铁死亡,从而对MASH产生治疗作用。这些结果表明,SGXZ是治疗MASH的重要汤,因此值得进一步研究。
----------微科盟精彩文章----------
综述 | Cell子刊:代谢组学在代谢功能障碍相关脂肪性肝病(MASLD)风险预测中的应用
科研 | 皖南医学院:甜叶菊根多糖调节肝脏代谢、胆汁酸和肠道菌群改善非酒精性脂肪肝(国人佳作)
如果需要原文pdf,请扫描文末二维码



获取此文献原文PDF、申请加入学术群,联系您所添加的任一微科盟组学老师即可,如未添加过微科盟组学老师,请联系组学老师47,无需重复添加。
请关注下方名片
随机文章
-
 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
- 东莞救护车出租转运服务流程解说】:遗体转运、担架车接送、跨省转院、护送服务、出院转院安捷重症接送
- 深圳打工人是不敢没有副业的
- 广州一阿姨家“火”了,都因为这种花,种1棵,长一片,引人侧目
- 东莞市石碣镇救护车转运服务平台:收费标准、长途护送、转院出院、跨省护送服务
- 中国5A景区:深圳观澜湖休闲旅游区
- 春节 • 悦玩港澳4日丨广州南进香港/深圳北出,珠澳港3晚品质4钻,高品纯玩不进不推不加点纯净粤港澳游 福建往返
- 劳动法条汇编--广州市南沙区人民政府办公室关于印发广州市南沙区见义勇为人员奖励和保障实施细则的通知
- 对照深圳最新的试卷看看新的思路
- 东莞市粤华学校2026年春季小初高插班生招生简章!
- 广州白鹅潭CBD江景甲级写字楼丨广州国际医药港·健康方舟
