具体而言,研究团队通过衣霉素和二硫苏糖醇诱导植物产生内质网胁迫,敏锐地捕捉到了一个关键现象:植物体内的m6A甲基化酶表达显著升高,全局m6A水平随之增加。遗传学分析进一步证实,那些m6A缺陷的突变体对胁迫更加敏感,而m6A积累增加的突变体则耐受性更强。
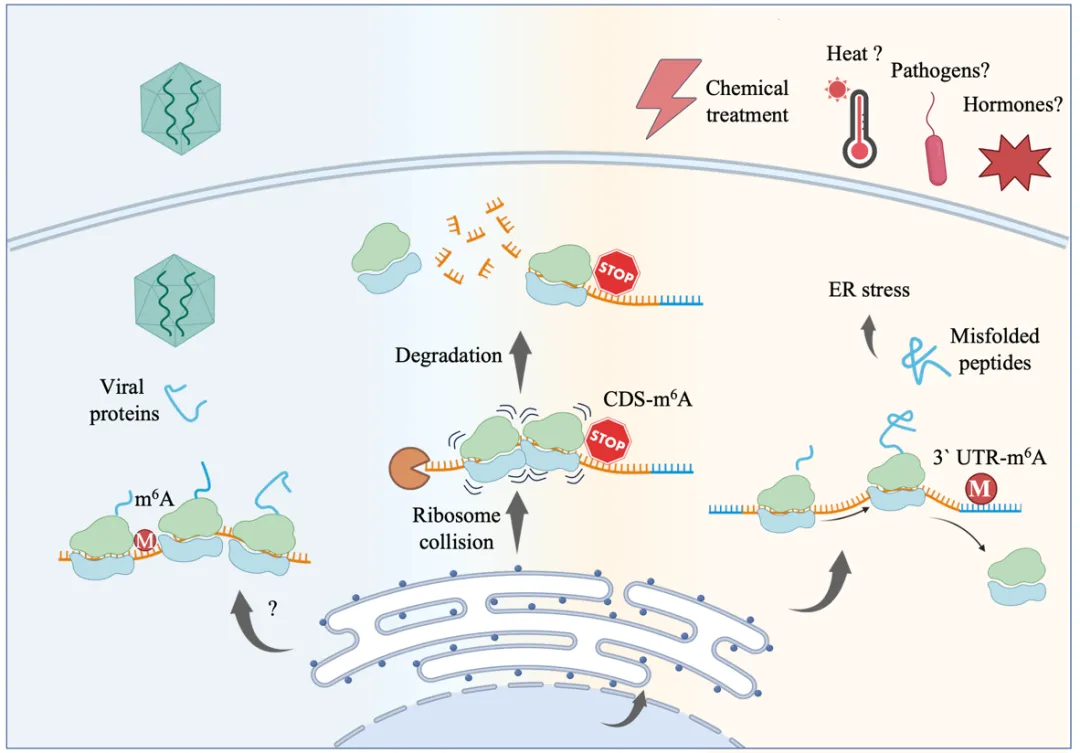
值得注意的是,该研究厘清了m6A在不同位置的“分工”:虽然植物体内大部分m6A修饰发生在RNA的“尾巴”区域(3’UTR),但真正在内质网胁迫中发挥关键减压作用的是位于编码区(CDS)的m6A。
那么,m6A究竟是如何发挥作用的?
研究团队结合了核糖体印记测序、m6A测序以及降解组测序进行联合分析。他们发现:m6A修饰并非随机分布,而是特异性地出现在RNA的编码区。当核糖体在翻译过程中遇到这些位于编码区的m6A修饰时,会发生停滞甚至碰撞,这种异常信号随即触发了RNA的降解程序。
这是一种极其精准的“共翻译RNA降解”机制——还没等这条RNA翻译成完整的蛋白质,它就被直接拆解销毁了。这就好比工厂流水线发现某个环节压力过大,直接剪断原材料输送带,从源头上切断蛋白质的合成,极大减轻了内质网的负担。
更有趣的是,这套精密的监控系统不仅在植物自身抗压中发挥作用,还成为了抵御病毒的“天然防线”。面对种类最丰富且对全球作物产量危害最严重的植物病毒类群之一——双生病毒,植物的m6A系统能利用其结构紧凑、编码区极易暴露的特点,精准识别并降解病毒RNA。研究发现,在m6A水平降低的突变体中,病毒RNA复制显著增强,植株对病毒侵染也表现出更高敏感性。这意味着植物巧妙地利用了同一套机制,既应对了非生物胁迫,又防御了生物胁迫。








 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
